Chromosomowa teoria dziedziczenia
W 1902 roku Sutton I Boveri skorelowali teorię dziedziczenia Mendla z zachowaniem się chromosomów w czasie mitozy i mejozy. Główne wnioski to: Geny zlokalizowane są na chromosomach, a chromosomy występują w parach. Chromosomy homologiczne są rozdzielane w czasie mejozy. Produkty mejozy zawierają jeden z chromosomów homologicznych, a w procesie zapłodnienia dochodzi do przywrócenia wyjściowej liczby chromosomów.
Thomas Morgan (1911), amerykański naukowiec, prowadził swoje prace oparte na badaniach eksperymentalnych i wyliczeniach matematycznych na muszkach owocowych (Drosophila melanogaster) w pierwszej połowie XX wieku. Muszka owocowa jako organizm modelowy ma wiele zalet. Należą do nich m.in: małe rozmiary, wyraźne różnice między samicami a samcami (dysmorfizm płciowy), dobrze wyrażone cechy zależne od pojedynczych genów, łatwość hodowli, mała liczba chromosomów (3 autosomy + 1 chromosom błci), znaczna płodność i krótki cykl życiowy.
Główne założenia chromosomowej teorii dziedziczności Morgana to:
Geny zlokalizowane są w chromosomach jąder komórkowych (chromosomowa teoria dziedziczności). Geny są powielane (ulegają replikacji), a każdy gen zajmuje na chromosomie ściśle określone miejsce (locus). Allele tego samego genu znajdują się w chromosomach homologicznych (od ojca i od matki) dokładnie naprzeciw siebie, mają to samo lokus. Allele różnych genów zajmują inne miejsca. Geny komórek potomnych są identyczne z genami rodzicielskimi.

Geny ułożone są w chromosomach liniowo w ściśle określonym porządku i kolejności.
Geny leżące na tym samym chromosomie są to geny sprzężone, a na odrębnych chromosomach- niesprzężone. Niezależnie dziedziczone są tylko te geny, które leżą w różnych chromosomach i są niesprzężone. Geny zlokalizowane w tych samych chromosomach tworzą tzw. grupę sprzężenia. Oznacza to, że dziedziczą się razem, co jest niegodne z drugim prawem Mendla. Geny sprzężone dziedziczone są razem, ale nie zawsze całkowicie. W czasie mejozy każdy z chromosomów homologicznych podlega losowej segregacji do jednej z komórek.
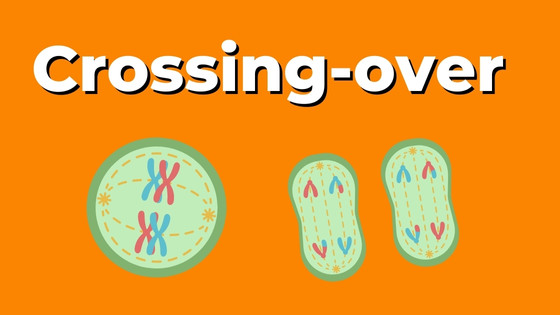
Rozprzężenie genów zachodzi w wyniku procesu crossing-over (c.o). Crossing-over to zjawisko, które ma miejsce w gametach na początku mejozy. Polega na wymianie odcinków chromatyd pomiędzy chromosomami homologicznymi. Między chromatydami tworzą się połączenia tzw. chiazmy. Ulegają one następnie rozerwaniu i następują wymiana odcinków chromosomów. Częstość crossing-over jest uwarunkowana położeniem genów w chromosomie. Jeżeli geny leżą blisko siebie, częstość crossing-over jest niewielka. W wyniku c.o powstają nowe kombinacje alleli. W wyniku c.o. powstają 4 klasy osobników: 2 mają układ cech taki sam jak rodzice (są to najbardziej liczne klasy), a 2 są rekombinantami-mają inny niż rodzice układ genów na chromosomie.
Czym większa odległość między dwoma genami na chromosomie, tym częstość występowania crossing-over jest większa. Na podstawie częstości crossing-over można określić położenie genów w chromosomach I sporządzić mapy chromosomów (ustalić położenie genów na chromosomie).
Wyniki krzyżówek genetycznych zależą od tego czy geny położone są na jednym czy na innych chromosomach.
Geny niesprzężone – co to takiego?
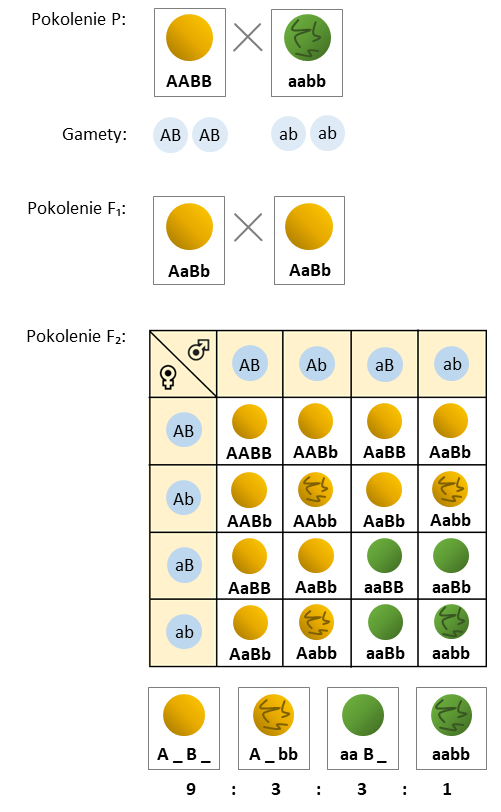
- Geny niesprzężone przechodzą do gamet niezależnie od siebie, zgodnie z II prawem Mendla. Stosunek fenotypów w F2 przy krzyżowaniu osobników o 2 różnych cechach to 9:3:3:1
Geny sprzężone – co to takiego?
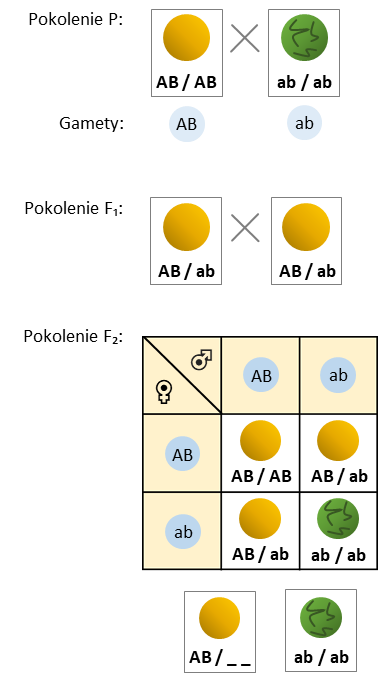
- Geny sprzężone – jeśli allele 2 genów znajdują się na tym samym chromosomie to przechodzą do gamet razem. Oznacza to, że dziedziczą się niezgodnie z drugim prawem Mendla. Przy założeniu, że barwa I struktura grochu znajdują się na jednym chromosomie, stosunek fenotypów w F2 wynosiłby 3:1
Mapowanie genów – odległość między genami na chromosomie
Odległość między genami określa się na podstawie częstości zachodzenia crossing-over między dwoma genami sprzężonymi AaBb aabb. W tym celu wykonuje się dwugenową krzyżówkę testową pomiędzy podwójną homozygotą recesywną a badanym osobnikiem. Procent uzyskanych rekombinantów określany na podstawie cech fenotypowych oznacza procent zachodzących procesów crossing-over. Odległość między genami określa się przeliczając liczbę uzyskanych rekombinantów na procenty. Jednostką odległości między genami jest 1 centimorgan (1 jednostka makowa) 1 j.m. odpowiada ona jednemu rekombinantów z krzyżówki wstecznej podwójnej heterozygoty.