Strategie absorpcji węgla przez rośliny i fotooddychanie
Fotosynteza to proces wytwarzania związków organicznych z CO2 przy udziale energii świetlnej, charakterystyczny dla fotoautotrofów. Fotosyntezę dzieli się na dwa główne etapy – fazę jasną, zachodzącą przy udziale światła i fazę ciemną, niezależną bezpośrednio od światła.
Fotosynteza – charakterystyka faz
Faza jasna fotosyntezy służy wytworzeniu siły asymilacyjnej – ATP i NADPH – niezbędnej do przeprowadzenia fazy ciemnej. Zachodzi w granach chloroplastów. Faza ciemna natomiast polega na asymilacji CO2 kosztem wytworzonej w fazie jasnej energii i zachodzi w stromie chloroplastów. Fazę ciemną dzieli się na trzy etapy – karboksylację, redukcję oraz regenerację. Faza ciemna może przebiegać w różny sposób u różnych gatunków roślin.
Poniżej przedstawiamy specyfikę i porównanie rośliny C3 i C4.
Rośliny C3
Większość roślin to rośliny typu C3. U tych roślin faza ciemna obejmuje cykl Calvina, a jego pierwszy etap – karboksylacja – polega na przyłączeniu CO2 do pięciowęglowego RuBP – rybulozo-1,5-bifosforanu, co katalizowane jest przez enzym Rubisco. Powstaje sześciowęglowa cząsteczka, która niezwłocznie rozpada się na dwie trójwęglowe cząsteczki PGA – kwasu 3-fosfoglicerynowego. Stąd właśnie nazwa tej grupy – rośliny C3. W fazie redukcji następuje aktywacja, a następnie redukcja PGA do PGAL (aldehydu 3-fosfoglicerynowego) przy udziale NADPH i ATP. Następnie ma miejsce regeneracja, czyli odtworzenie pięciowęglowego akceptora CO2.

Fotooddychanie
Rośliny w klimacie tropikalnym w ciągu dnia są narażone na wysokie temperatury. W związku z tym przymykają szparki, aby ograniczyć transpirację. Wiąże się to jednak z odcięciem dostępu do CO2, co prowadzić może do fotooddychania. Jest to proces oksydacyjnego zużywania tlenu, podczas którego wytwarzany jest CO2. Zużywane ATP, a RuBP ulega oksydacji, przez co zmniejsza się wydajność fotosyntezy. Wykształciły jednak mechanizmy, minimalizujące proces fotooddychania poprzez zwiększenie dostępności CO2 dla enzymu Rubisco.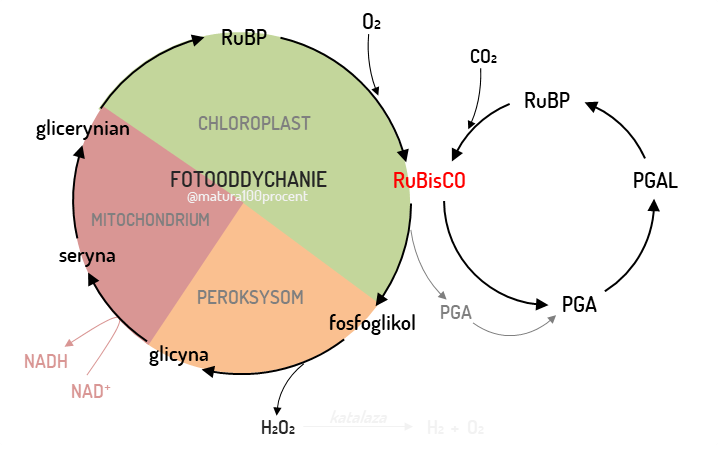
>> Chcesz dobrze zdać maturę z biologii? Zobacz ebook Botanika
Rośliny C4
Rośliny klimatu tropikalnego, takie jak kukurydza czy trzcina cukrowa posiadają układ wieńcowy, otaczający wiązki przewodzące. Dzięki niemu mogą sprawniej wykorzystywać CO2. Układ wieńcowy składa się z wewnętrznej warstwy pochwy wokółwiązkowej, w której zachodzi cykl Calvina, oraz mezofilu na zewnątrz, gdzie pierwotnie wiązany jest CO2. CO2 wiązany jest do fosfoenolopirogronianu (PEP). Powstaje czterowęglowy szczawiooctan, stąd nazwa tej grupy – rośliny C4. Szczawiooctan redukowany jest do jabłczanu, który trafia do pochwy wokółwiązkowej, gdzie ulega dekarboksylacji, dając pirogronian oraz CO2. CO2 jest gromadzony w komórkach pochwy wokółwiązkowej i wykorzystywany dalej w cyklu Calvina.

Rośliny CAM
Sukulenty wykształciły inny mechanizm minimalizujący fotooddychanie. CO2 przyłączany jest do PEP, dając szczawiooctan, który następnie ulega redukcji do jabłczanu. Jabłczan gromadzony jest w wakuoli, powodując jej zakwaszenie. Kiedy w ciągu dnia szparki są zamknięte i roślina nie pobiera CO2, wykorzystywany jest zgromadzony w wakuolach jabłczan. Przenoszony jest do cytozolu, gdzie ulega dekarboksylacji do CO2, który wykorzystywany jest dalej w cyklu Calvina, dzięki zachodzącej wówczas fazie jasnej, w której produkowana jest siła asymilacyjna
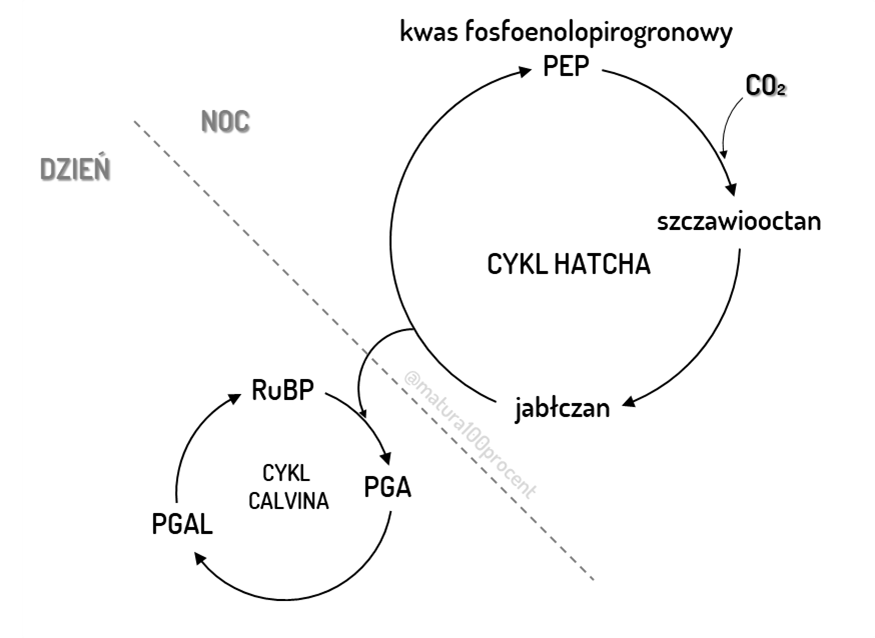
Porównanie rośliny C3, rośliny C4 i rośliny CAM
U roślin C3 występuje jeden akceptor CO2 – RuBP, a u rośliny C4 i CAM dwa – pierwotny PEP i wtórny RuBP. Produktem wiązania CO2 u roślin C3 jest sześciowęglowa cząsteczka, która rozpada się na dwie trójwęglowe cząsteczki PGA. Produktem pierwotnego wiązania CO2 u rośliny C4 i CAM jest czterowęglowy szczawiooctan. U roślin CAM asymilacja CO2 i cykl Calvina są rozdzielone w czasie. Asymilacja zachodzi w nocy, kiedy aparaty szparkowe są otwarte, a cykl Calvina w dzień, kiedy aparaty szparkowe są przymknięte ze względu na wysoką temperaturę. U roślin C4 faza ciemna rozdzielona jest w przestrzeni – początkowe przyłączenie CO2 zachodzi w komórkach mezofilu, a cykl Calvina zachodzi w komórkach pochwy wokółwiązkowej.
Przypomnij sobie też, na czym polega oddychanie tlenowe i jak wygląda budowa pierwotna łodygi.
Sprawdź kursy maturalne, które oferujemy!